
Chronic Low-Dose Radioactive Exposure:
False Alarm or Public Health Hazard ?
Wolfgang Koehnlein. Direktor of the Institute for Radiation Biology,
University of Muenster,
48129 Muenster, Robert-Koch-Strasse 43, Germany
Rudi Naussbaum. Portland State University, Portland OR 97205-0751/USA
For more than one hundred years, ionizing radiation and its biological effects have been a subject of scientific investigation. In these investigations, Dose-Effect-Relation is a most important correlation in proving causality between absorbed radiation dose and observed effect; that is, where effect is observed to increase with dose of a physical or chemical agent, then the agent can be considered as a cause. In Dose-Effect Rate research, not only is investigation of deterministic radiation effects from high radiation exposure important, the stochastic effects from low dose exposure are also important. The former high radiation exposure, through cancer induction, can affect an exposed person, the latter, low dose exposure, through genetic mutation, can affect the person's descendants.
It is therefore understandable, that the course of dose-effect should become a very significant point of discussion. The question that concerns us, then, is whether there exists a safe-dose threshold, a low-dose or low-dose-rate of low LET radiation, below which no detrimental effect can occur, either cancer morbidity or genetic mutations in an exposed population or their offspring.
Since 1927 the permitted dose levels of radiation exposure have been reduced continuously. Should the idea of a safe-dose and safe-dose rate become the prevailing scientific opinion, in spite of research indicating otherwise, then intended and un-intended exposures to raditation would increase above the present level, and premature cancers and additional mutations would occur world wide.
The terms 'safe dose' and 'safe dose rate', require clarification. In ionizing radiation, a safe dose and safe-dose rate means that all exposed persons remain unharmed during and after the exposure. No one will later on suffer from a radiation induced cancer or die prematurely. An unsafe dose or dose-rate, on the other hand, means that no one is safe during exposure, and that afterwards, a certain fraction of those exposed will suffer from radiation induced cancer and die prematurely. In the literature of this debate, there are numerous statements by experts and expert commissions claiming that there is an actual 'safe-threshold dose' of low LET radiation. While others consider that the question of a safe-dose threshold cannot yet be answered, my research indicates that enough data of exposed populations exists to show that there is no safe dose, no safe-dose rate, nor a safe dose threshold. Corresponding data with model systems similarly show that there is no safe dose threshold [8, 9]. I will confine this study, however, to observations made with exposed groups of humans, for the following reasons:
(1) Transfer of the results from the model system to human beings is unnecessary and less appropriate.[10]
(2) Numerous accepted investigations are available for exposed human beings in the low dose range relevant for radiation protection.[11, 12, 13, 14, 15, 16.]
These are the starting points of my consideration:
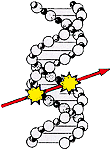
(1) The dose from low-LET ionizing radiation is delivered by high speed electrons (compton- and photoelectrons) traveling through human cells and creating primary ionization tracks. One such track is the least possible disturbance which can occur at the cellular level. A "high dose" means many tracks per cell; "low dose" means few tracks per cell; "low dose rate" means few tracks per cell per unit time. Whenever there is any dose at all, it means some cells and cell-nuclei are being transversed by tracks.
(2) Radiation induced carcinogenic alterations mean alterations in the genetic material of the cell, in the DNA. Cancer initiation is a unicellular process following the rules of chance. Every track, without the help from any other track, has a chance of inducing cancer by creating such alterations. The energy deposited during a primary interaction is many times the chemical binding energy in organic molecules.
(3) This implies that there can never be any safe dose or dose rate. However, if every carcinogenic alteration induced by tracks were successfully and invariably "undone" by repair processes, then there would be an inherently safe dose and dose rate. The key question is: Does repair of carcinogenic injuries operate flawlessly, when dose is sufficiently low and slow?
(4) If a radiation dose is received in the time frame required per repair, and if repair operates flawlessly and leaves no carcinogenic damage, then the net effect of that radiation dose toward cancer-production is obviously zero, by definition. It follows that many such small doses, with the corresponding repair times in between, could be absorbed without increasing the cancer rate.
(5) Human epidemiological evidence shows that repair fails to prevent radiation induced cancer, even at doses where the repair system has to deal with only one or a few tracks at a time and even at dose rates which allow ample time for repair before arrival of additional tracks (damage). Such evidence is proof, by any reasonable standard, that there is no dose or dose rate which is safe.
Before discussing several examples I will comment on the number of primary ionisation tracks traversing a cell nucleus at a given dose. In order to find out whether there is any safe dose it is appropriate to convert a given dose in fractions of Sievert into number of tracks through the cell nucleus, since the smallest possible dose is not a fraction of a Gray, but a single traversal of an ionizing track through the cell nucleus. As we know, the energy of x- and g-rays is deposited in biological material via photo- and Compton - electrons. One can, therefore, use the calculations of Paretzke and a recursion method [17] to convert the energy of a x- or g-ray into electrons and their energy distribution. Thus, it is possible to transfer the original photon energy to electrons and calculate their summarized range [18, 19]. With help of the relation 1cGy - 6,24. 1010 keV/g we can now determine how many photons of a given energy are required to deposit a dose of 1 cGy. With this information the total sum of the electron tracks is obtained. Since the average dimensions of a mammalian cell and its nucleus are known [20], we arrive at the number of nuclear traversals per dose unit for x- and g-radiation of different origin.
Table 1: Tissue Dose in Centi-Gray when the Average Track-Rate per Cell Nucleus is One
Origin of Electron Packets Average Nuclear Traversals per Nucleus in Delivery of 1 cGy Dose in Centi-Gray when Average Track-Rate per Nucleus is one 30 keV X-rays 1.3379 0.747 cGy 250 keV X-rays 2.3 0.434 cGy 596 keV gammas Radium-226 2.937 0.340 cGy 662 keV gammas Cesium-137 3.156 0.317 cGy 1608 keV gammas A-Bomb 5.407 0.185 cGy
We know from numerous experiments with model systems, that enzymatic repair processes are working without impairment even at doses of a few Grays [21, 22, 23, 24, 25]. It has also been confirmed, repeatedly, in studies with human cells in vitro, that whatever repair is achieved is complete within 6 hours or less, even after doses of several Grays [26, 27, 28, 29]. There is also confirmed information on the number and type of DNA lesions (Table 2). There are, however, numerous references in the literature that certain DNA-lesions are not repaired or misrepaired. To give some examples: The UNSCEAR-Report 1986 (30] states the following on repaired, unrepaired and misrepaired carcinogenic lesions induced by radiation:
Table 2: Number of lesions induced in the DNA of a mammalian cell
per D(37) of absorbed radiation energy
DNA-Single Strand breaks 1000 DNA-Double Strand breaks 50 Base Damage 200 Protein-DNA Crosslinks 150 Bulky Lesions 450
"The error-free repair of the DNA, the most likely target involved, leaves some fractions of the damage unrepaired and the error-prone repair may produce misrepaired sequences in the DNA-structure."
Kellerer describes a type of radiation induced DNA damage which would be difficult to repair.[22] "A simple example would be neighboring single-strand breaks in complementary strands of DNA, which interfere with excision repair."
This is confirmed by Feinendegen et al.[27], reporting that in radiated cells not all double-strand breaks are fully repaired. With the material discussed so far, we are able to examine whether any safe dose does exist.
Consider the following scenario: The repair processes work flawlessly up to a certain dose of a few cSv (100 cSv - 1 Sv). A number of individuals are exposed to a small dose on Monday. All induced lesions in the DNA are flawlessly repaired within a few hours. No increase of cancer risk results from this exposure. On Tuesday there is the next exposure with the same small dose. Since the repair systems are working error free, there is no increase in cancer risk after the first two doses. On the following days further dose fractions are given, and so on. In this scenario the individuals could accumulate rather high doses in many small dose fractions. There would be no increased cancer risk detectable, however, in a long term follow-up, while it is known that the total dose given at once will increase the cancer risk we could still conclude that the given dose fractions are harmless and that a threshold dose is real. If, however, the long term follow-up studies reveal increased cancer incidence in the exposed population even though they were exposed to small dose fractions only, the the conclusion of an error-prone repair system even at lose doses would be obvious. It also follows that the idea of a safe threshold dose is incorrect.
For a number of carefully carried out epidemiological studies of exposed persons which are accepted in the scientific literature, the dose fractions or doses respectively and the derived number of tracks per cell nucleus per exposure are compiled in the table. In all nine studies a significant increase in cancer incidence has been observed in the exposed population.
Table 3: Average Tracks per Cell Nucleus from each Exposure in Nine Human Studies
Nr. Study Organdose[cSv]/-Exposure Tracks per Cell Nucleus at 1 cSv Average Number of Tracks per Cell Nucleus from each Exposure 1 Nova Scotia Fluoroscopy -[ 13] 7,5 1,3378 10,0335 Rounded : 10 2 Israeli Scalp-Irradiation (Autors�revised est.) 7,5
9.51.3378 10,0335 Rounded : 10
12,0402 Rounded : 123 Massachusetts Fluoroscopy 4,6 1,3378 6,1539
Rounded : 64 Canadian Fluoroscopy 4,6 1,3378 6,1539
Rounded : 65 Stewart In-Utero Series 0,5 1,3378 0,6689~ one;
51% with on track6 MacMahon In-Utero Series 0,9 1,3378 1,2040~ one 7 British Luminizers [15] 0,1 2,9370 0,293~ one;
75% with no track8 Harvey Twins In-Utero Series 1,0 1,3378 1,3378~ one 9 Israeli Breast-Cancer in Scalp-Irradiation Study -- [11] 1,6 1,3378 2,140~ two
1. See Ref. 13.
2. See Ref. 11.
3. Boice JD, Monson, RR, Rosenstein M. Cancer mortality in women after repeated fluoroscopic examinations of the chest, Journal of the National Cancer Institute 66, 863-867, 1981;
4. Miller AB, Howe GR, Sherman GJ, Lindsay JP, Yaffe MJ, Dinner PJ, Risch HA, Preston DL. Mortality from breast cancer after irradiation during fluoroscopic examinations in patients being treaded for tuberculo-sis, New England Journal of Medicine 321, 1285-1289, 1989.
5. Gilman EA, Kneale GW, Know EG, Stewart AM. Pregnancy X-rays and childhood cancers: Effects of exposure age and radiation dose. Journal Radiol. Protection 8, 3-8, 1988.
6. MacMahon B. Prenatal X-ray exposure and childhood cancer. Journal of the National Cancer Insitute 28, 1173-1191, 1962.
7. See Ref. 15.
8. Harvey EB, Boice JD Jr., Honeyman M, Flannery JT. Prenatal X-ray exposure and childhood cancer in twins, New England Journal of Med. 312, 541-545, 1985.
9. See Ref. 11. These studies show that the following doses cannot be regarded as safe with respect to cancer induction.
We can therefore conclude that whenever an ionizing track traverses a nucleus of a mammalian cell, there is always a chance that it will cause a carcinogenic lesion and that the lesion will be repaired, inherently unrepairable or misrepaired. In short, there is an inherent failure rate in the repair system. For the essential stochastic end points of radiation damage (cancer induction and mutation) the idea of a safe threshold dose and of a safe dose range must be rejected. According to the present scientific knowledge from the various proposed dose-effect relationships shown in the figure 6, only the linear (3), or the supralinear (4) is consistent with scientific evidence on human data.
The supporters of threshold doses and even hormetic effects will certainly argue that there are many studies where there was no radiation effect found by the authors. However these studies are unsuited to deciding whether there is a threshold dose or not. This has also been admitted by the authors. The finding of "no effect" can never be an argument for or against a safe dose. Very often the follow up periods were too short, the size of the cohorts too small and the confounding factors were not taken properly into account. A group of 12 independent scientists (physicians and epidemiologists) (31), sponsored by "Physicians for Social Responsibility" have critically reviewed 124 epidemiological studies supported or financed by the U.S. DoE and by the British Government. Their critical report concludes that:
1. ...the DoE's epidemiological program is seriously flawed...
2. ...the problems and flaws evident in many investigations are precisely those which tend to produce false negative results...
It is, therefore, unsurprising that a large number of epidemiological mortality studies show no significant correlation between cancer induction and low dose radiation exposure.
Nevertheless, arguing from scientific facts and independent epidemiological studies clearly show that the repair systems of the mammalian cell is never 100% exact and that there is no harmless dose threshold. That this has still not been accepted by most national and international commissions suggests that official estimates are no longer a scientific process but rather a political one.
Literature
1.Gilbert ES, Omohundro E, Buchanan JA et al. Mortality of workers at the Hanford site: 1945-1986, Health Phys. 64, 577 - 590,1993
2.Cardis E, Gilbert ES, Carpenter L. et al. Direct estimates of cancer mortality due to low doses of ionising radiation; an international study. Lancet 344,1039 - 1043,1994
3.Goldman M. Cancer risk of low-level exposure, Science 271 1821 - 1822,1996
4.BEIR V. National research council. Health effects of exposure to low levels of ionising radiation. Washington, DC: National Academy Press 1990
5.Land CE. Estimating cancer risks from low-doses of ionising radiation. Science 209,1197 - 1203,1980
6.Nussbaum RH, Koehnlein W, Belsey E. Die neueste Krebsstatistik der Hiroshima-Nagasaki-Ueberleben- den, Med. Kiln. 86,90 - 108,1991
7.Nussbaum RH, Koehnlein W. Inconsistencies and open questions regarding low-dose health effects of ionising radiation, Environ. Health Persp. 102,656-667,1994
8.Grosovski AJ, Little JB: Evidence for linear response for the induction of mutations in human cells by x- ray exposures below 10 rads. Proc. Nati. Acad. Sci. USA 82,2092 - 2095,1986
9.Waldren C, Correl L, Sognier A, Puck 71: Measurement of low levels of x-ray mutagenesis in relation to human disease. Proc. Nati. Acad. Sci, USA 83,4839 - 48441986
10.Fry RJM. Relevance of animal studies to the human experience. ln:Boice JD Jr., Fraumeni JD Jr. Radiation carcinogenesis and biological significance. Raven Press New York 1984,337 - 346
11.Modan B, Alfandry E, Cherit A, Katz L. Increased risk breast cancer after low dose irradiation. In: Lancet Nr. 8639 Bd. 1,629 - 631,1989
12.Sorahan T, Roberts PJ. Childhood cancer and parental exposure to ionising radiation: Preliminary findings from the Oxford survey of childhood cancers, Am. J. Ind. Med. 23, 343 - 354,1993
13.Myrden JA, Hiltz JE. Breast cancer following multiple fluoroscopies during artificial pneumothorax treatment of pulmonary tuberculosis, Canadian Medical Ass. J. 100,1032 - 1034,1969
14.Bithell JF, Stiller CA. A new calculation of the radiogenic risk of obstetric X-raying. Stat. Medicine 7, 857-864,1988
15.Baverstock KF, Papworth DG, The UK radium luminizer survey. Brit. J. of Red. Supplemental BIR Report2l,71 -76,1987
16.for additional references see for example: Nussbaum RH Koehnlein W. Health consequences of exposures to ionising radiation from external and internal sources: challenges to radiation protection standards and biomedical research, Medicine & Global Survival 2,198 - 213,1995
17.Paretzke HG. Radiation track structure theory. Chapter 3, p 89-169 In: Freeman GR (Ed.) Kinetics of nonhomogeneous Processes. New York: John Wiley and Sons, 1987.
18 Evans RD. Stopping of electrons by thick absorbers, Chapter 21, p 611-631 in The Atomic Nucleus. New York, McGraw Hill, 1955
19.Hutchinson F. Formation of two double-strand breaks in the same DNA molecule by a single high- energy photon or ionising particle, Int. J. Radiat. Biol. 70,505-512,1996
20.Brackenbusch LW, Braby LA. Microdosimetric basis for exposure limits, Health Phys. 55,251 - 255, 1988
21.Virsik RP, Blohm R, Herman KP, Modler M und Harder D. Proceedings of the eighth symposium of microdosimetry, p.409 - 422 (Euratom 8395, Luxemburg) 1982
22 Kellerer AM. Models of cellular radiation action, chapter 7, 305 - 375, in: Freeman GR (ed.) Kinetics of non homogeneous processes. New York, John Wiley & Sons
23.Frankenberg D, Goodhead DT, Frankenberg-Schwager M, Harbich R Bance DA und Wilkinson RE. Effectiveness of 1,5 keV aluminium and 0,3 keV carbon K characteristics X-rays at inducing DNA double- strand breaks in yeast cells. lnternat.J.of Radiat. Biol. 50,727-741,1986
24.Frankenberg-Schwager M, Frankenberg D, Harbich R, Adamczyk R: A comparative study of rejoining of DNA double-strand breaks in yeast irradiated with 3.5 MeV a-particles or with 30 MeV electrons. Int. J. Radiat. Biol. 57,1151 - 1168,1990
25.Peak MJ, Wang L, Hill CK, Peak JG. Comparison of repair of DNA double-strand breaks caused by neutron or gamma radiation in cultured human cells. Int. J. Radiat. Biol. 60,891 - 8981991
26.Bender MA. Significance of chromosome abnormalities, p. 281 - 289 in Radiat. Carcinogenesis: Epidemiology and biological significance, eds. Boice JD and Fraumeni JF, Raven Press, New York, 1984
27.Feinendegen LE, Victor PB, Booz J, Muhlensiepen H. Biochemical and cellular mechanisms of low- dose effects. Int. J. of Radiat. Biol. 53,23 - 37,1988
28.Burns FJ, Sargent EV. The induction and repair of DNA breaks in rat epidermis irradiated with elec- trons. Radiat. Res., 87,137-1441981
29.Frankenberg-Schwager M. Review of repair kinetics for DNA damage induced in eukaryotic cells in vitro by ionising radiation. Radiother. Oncol. 14,307 - 320,1989
30.UNSCEAR or United Nations Scientific Committee on the Effects of Atomic Radiation, 1986. Genetic and somatic effects of ionising radiation, New York, United Nations
31.Geiger JH, Rush D, Michaels D, Baker DB, Cobb J, Fischer E, Goldstein A, Kahn HS, Kirsch JL, Landrigan PJ, Mauss E und Mclean DE: Dead reckoning: A critical review of the Department of Energy's Epidemiological research. Washington DC: Physicians for Social Responsibility 1992
![[Button]](/images/greendot.jpg)
![[Button]](/images/arc.jpg)
|
![[Button]](/images/leaf.gif)
|